Thread by Erez Lieberman Aiden
Thread
Excited about a new collaboration between the Rowland, Onuchic, Di Pierro, and @theaidenlab labs and @thednazoo which was led by @odudcha Ahmed Elbatch, Claire Hoencamp and Sumitabha Brahmachari, appearing in @ScienceMagazine today. A tweetorial! 
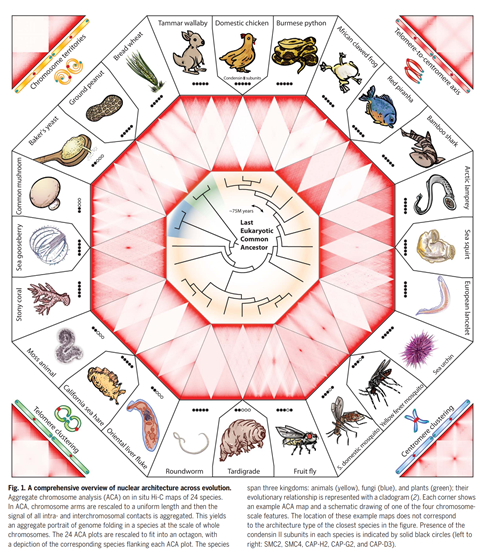
The team @thednazoo, led by @odudcha, spanning institutions ranging from @stanford to CTBP to @seaworld, created dozens of chromosome-length & diploid genome assemblies across the tree of eukaryotic life. We found that chromosomes inside nuclei fold in one of two overall designs. 

One, seen in mushrooms & mosquitoes, is associated w/ telomere2telomere contacts, centromere2centromere contacts,& a telomere-to-centromere axis. The other, seen in humans & sea hares, has chr territories. Types seem to switch alongside evolutionary changes to condensin II.
So what happens if you disrupt Condensin II in the lab? In gorgeous experiments, Claire Hoencamp of the Rowland lab showed that if you disrupt condensin II in human cells, the cells change from Type II to Type I. But only if you let them go through mitosis after disruption. 

These experiments were terrific and things started to become clear.
Sumitabha Brahmachari in the Onuchic lab used physical simulations to show that this can be explained the tendency of condensin II, a loop extrusion protein, to linearly compact chromosomes. 

Interestingly, evolution demonstrated this point in a different way: the Chinese muntjac has 46 chromosomes, the closely-related Indian muntjac only 6, much much longer chromosomes. The former is Type II; the latter, type I. 

We think that the tendency of condensin II to form shorter, stiffer chromosomes during metaphase makes it harder for centromeres to find each other and form territories. Instead you get a Type I architecture which resembles anaphase in many ways. 

A final note: our paper introduces the new Phaser module for 3D-DNA. If you have an unphased genome, SNP calls, and adequate Hi-C data (30X-60X in human), you can now create diploid assemblies easily. 

But in some species, like drosophila, homologs are closely paired and this strategy does not work. This is a great example of how changes in nuclear architecture can necessitate different genome assembly strategies across the tree of life. 

This work is a fun example of how, using rapid genome assembly across numerous species, it's possible to get going doing genomics in any species you like, and to deploy the whole tree of life to study a problem. 

It was really wonderful to tackle these problems with a truly amazing team!
Read the paper at: science.sciencemag.org/content/372/6545/984.full
If you're interested in these topics, also check out Abdennur et al. on @biorxivpreprint from the Spitz and Mirny groups (on condensin disruption) and recent work on Hi-C phasing by Kronenberg et al @NatureComms and Garg et al @NatureBiotech